Mitochondria are double-membraned organelles responsible for integrating metabolic pathways for energy production and the coordination of cellular stress responses; namely, apoptosis, hypoxia and autophagy. Considered the powerhouses of the cell, mitochondria release chemical energy stored in glucose and other nutrients in a series of exothermic redox reactions. This is enabled by the production of high-energy electron carriers, NADH and FADH. The process of aerobic respiration initiates in the cytosol during glycolysis. Here, glucose is oxidised and split into two pyruvate molecules, producing two ATP and two NADH. Next, pyruvate is transported across both mitochondrial membranes into the luminal matrix space. The following decarboxylation step produces an additional NADH as pyruvate is converted in to acetyl-CoA; an entry-point intermediate within the circular metabolic pathway known as the Kreb’s cycle. The Kreb’s cycle does not directly generate ATP but instead produces GTP, FADH and three more NADH. The availability of reduced NADH and FADH molecules feeds the third stage of respiration, oxidative phosphorylation. The oxidation of NADH and FADH allows electrons to be transferred across a series of enzymatic complexes (I-IV) integrated within the inner mitochondrial membrane. The energy released in this electron transport chain (ETC) is used to pump protons from the matrix into the intermembrane space. This creates an electrochemical gradient across the inner membrane which is harnessed by the membrane-bound ATP synthase complex to drive ATP formation; a phenomenon called chemiosmosis. NADH oxidation occurs at ETC complex I and is responsible for the movement of ten protons into the intermembrane space. Notably, FADH oxidation only powers the transfer of six protons. This is because FADH-derived electrons enter the ETC downstream at complex II, bypassing the proton-pump activity of complex I. Components of ETC enzymatic complexes constitute useful mitochondrial marker proteins as their expression correlates with mitochondrial density and function. These include: NDUFV2, NDUFS1 (complex I); SDHA-D (complex II); BCS1L, UQCRC1 (complex III); COX2, COX7A2L, COXIV (complex IV); and ATP50, ATP5A (ATP synthase). Other marker proteins can be used to interrogate broader aspects of mitochondrial function. For example, in the matrix the chaperone HSP60 and protease CLPP both promote protein quality control, while outer-membrane bound receptors TOM20 and TOM70 facilitate mitochondrial protein import. The multi-functional anion channel VDAC is another marker of the outer mitochondrial membrane. Conformational changes in VDAC activate apoptosis by promoting mitochondrial outer-membrane permeabilisation and subsequent release of pro-apoptotic factors into the cytoplasm. Finally, several marker proteins inform researchers on mitochondrial dynamics and morphology. These include regulators of mitochondrial fusion (mitofusin1/2, OAP1) and fission (FIS1), as well as other modulators of organelle structure and membrane integrity (mitofilin, prohibitin). We offer a wide range of antibodies against mitochondrial markers including AIF antibodies, Galectin 3 antibodies, HIF-1 alpha antibodies, p53 antibodies, and HSP60 antibodies, that are validated across multiple applications and cover various host species, antibody types, conjugates, and formulations. Changes in the expression or localisation of these proteins can indicate alterations in mitochondrial activity which can be linked to neurodegenerative disorders, cancer or ageing.
Anti-HSP70 Antibody [C92F3A-5] (A304773) | |
200µg/$555 | |
| Description: | Mouse monoclonal [C92F3A-5] antibody to HSP70. |
| Applications: | WB, IHC, ICC/IF, ELISA, Flow Cytometry, FACS, IEM, Block/Inhibit, Antibody Microarray |
| Reactivity: | Human, Mouse, Rat, Bovine, C. elegans, Canine, Chicken, Drosophila, Carp, Guinea Pig, Hamster, Monkey, Porcine, Rabbit, Sheep |
| Conjugate: | Unconjugated |
![FACS - Anti-HSP70 Antibody [C92F3A-5] (A304773) - Antibodies.com](https://cdn.antibodies.com/image/catalog/304/A304773_1.png?profile=product_search)
Anti-HSP60 Antibody (A85438) | |
10µl - 100µl/$190 – $5451 Citation | |
| Description: | Chicken polyclonal antibody to HSP60. |
| Applications: | WB, ICC/IF, IHC |
| Reactivity: | Human, Rat, Mouse, Horse, Canine, Monkey, Porcine, Bovine |
| Conjugate: | Unconjugated |
Anti-HSP70 Antibody (A304824) | |
100µl/$520 | |
| Description: | Rabbit polyclonal antibody to HSP70. |
| Applications: | WB, IHC, ICC/IF, IP, ELISA |
| Reactivity: | Human, Mouse, Rat, Beluga, Bovine, Canine, Fish, Carp, Guinea Pig, Hamster, Monkey, Porcine, Sheep, Coral, Plant, Shark, L. amazonensis |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [LK1] (A305088) | |
200µg/$420 | |
| Description: | Mouse monoclonal [LK1] antibody to HSP60. |
| Applications: | WB, IHC, IP, ELISA, Flow Cytometry |
| Reactivity: | Human, Mouse, Rat, Bovine, Canine, Chicken, Drosophila, Guinea Pig, Hamster, Monkey, Porcine, Rabbit, Sheep, Xenopus, Plant, Silkworm |
| Conjugate: | Unconjugated |
![Immunocytochemistry/Immunofluorescence - Anti-HSP60 Antibody [LK1] (A305088) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305088_1.png?profile=product_search)
Anti-Tau Antibody [2E9] (A85416) | |
10µl - 100µl/$190 – $545 | |
| Description: | Mouse monoclonal [2E9] antibody to Tau. |
| Applications: | WB, ICC/IF, IHC |
| Reactivity: | Human, Horse, Cow, Porcine, Rat, Mouse |
| Conjugate: | Unconjugated |
Anti-HSP70 Antibody [BB70] (A305113) | |
200µg/$540 | |
| Description: | Mouse monoclonal [BB70] antibody to HSP70. |
| Applications: | WB, ICC/IF, IHC, IP |
| Reactivity: | Human, Mouse, Rat, Bovine, Sheep, Canine, Beluga, Fish, Guinea Pig, Porcine, Hamster, Rabbit, Chicken, Xenopus, Drosophila, Yeast |
| Conjugate: | Unconjugated |
Anti-p53 Antibody (A98895) | |
10µg - 100µg/$190 – $475 | |
| Description: | Rabbit polyclonal antibody to p53. |
| Applications: | WB, IHC, ELISA |
| Reactivity: | Human, Rat |
| Conjugate: | Unconjugated |
Anti-HSP70 Antibody [N27F3-4] (A305087) | |
200µg/$560 | |
| Description: | Mouse monoclonal [N27F3-4] antibody to HSP70. |
| Applications: | WB, IHC, ICC/IF, IP, Flow Cytometry, IEM |
| Reactivity: | Human, Mouse, Rat, Bovine, C. elegans, Beluga, Canine, Chicken, Drosophila, Fish, Guinea Pig, Hamster, Monkey, Porcine, Plant, Rabbit, Sheep, Xenopus |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [LK2] - BSA and Azide free (A252065) | |
100µg/$545 | |
| Description: | Mouse monoclonal [LK2] antibody to HSP60. |
| Applications: | IF, WB, IHC-P |
| Reactivity: | Human, Monkey, Mouse, Rat, Hamster, Guinea Pig, Chicken, E. coli, Borellia, Streptococcus pyogenes, Yersinia entercolitica, Salmonella typhimurium, Treponema hyodysenteriae, Treponema innocense, Trichinella spiralis, Helminths, Yeast, Spinach |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [LK2] (A248885) | |
100µg/$545 | |
| Description: | Mouse monoclonal [LK2] antibody to HSP60. |
| Applications: | IF, WB, IHC-P |
| Reactivity: | Human, Monkey, Mouse, Rat, Hamster, Guinea Pig, Chicken, E. coli, Borellia, Streptococcus pyogenes, Yersinia entercolitica, Salmonella typhimurium, Treponema hyodysenteriae, Treponema innocense, Trichinella spiralis, Helminths, Yeast, Spinach |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [LK2] (A305090) | |
200µg/$480 | |
| Description: | Mouse monoclonal [LK2] antibody to HSP60. |
| Applications: | WB, IHC, Flow Cytometry |
| Reactivity: | Human, Mouse, Rat, Bovine, Canine, Chicken, Guinea Pig, Hamster, Monkey, Porcine, Rabbit, Plant, Bacteria, Trichinella spiralis, Yeast, Insect, Fish |
| Conjugate: | Unconjugated |
Anti-Tau Antibody (A85414) | |
10µl - 100µl/$190 – $545 | |
| Description: | Chicken polyclonal antibody to Tau. |
| Applications: | WB, ICC/IF, IHC |
| Reactivity: | Human, Horse, Bovine, Porcine, Rat, Mouse |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [LK1] - BSA and Azide free (A252063) | |
100µg/$545 | |
| Description: | Mouse monoclonal [LK1] antibody to HSP60. |
| Applications: | Flow Cytometry, IF, WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken, Xenopus laevis, Drosophila |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [HSPD1/780] - BSA and Azide free (A252067) | |
100µg/$545 | |
| Description: | Mouse monoclonal [HSPD1/780] antibody to HSP60. |
| Applications: | Flow Cytometry, IF, WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken |
| Conjugate: | Unconjugated |
Anti-Galectin 3 Antibody (A94546) | |
10µg - 100µg/$190 – $475 | |
| Description: | Rabbit polyclonal antibody to Galectin 3. |
| Applications: | WB, IHC, IF, ELISA |
| Reactivity: | Human, Mouse, Rat |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [LK1] (A248883) | |
100µg/$545 | |
| Description: | Mouse monoclonal [LK1] antibody to HSP60. |
| Applications: | Flow Cytometry, IF, WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken, Xenopus laevis, Drosophila |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [HSPD1/780] (A248887) | |
100µg/$545 | |
| Description: | Mouse monoclonal [HSPD1/780] antibody to HSP60. |
| Applications: | Flow Cytometry, IF, WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [SPM253] - BSA and Azide free (A252064) | |
100µg/$545 | |
| Description: | Mouse monoclonal [SPM253] antibody to HSP60. |
| Applications: | Flow Cytometry, IF, WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken, Xenopus laevis, Drosophila |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody (A85437) | |
10µl - 100µl/$190 – $545 | |
| Description: | Rabbit polyclonal antibody to HSP60. |
| Applications: | WB, ICC/IF, IHC |
| Reactivity: | Human, Rat, Mouse, Bovine, Porcine, Horse, Canine, Monkey |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [SPM253] (A248884) | |
100µg/$545 | |
| Description: | Mouse monoclonal [SPM253] antibody to HSP60. |
| Applications: | Flow Cytometry, IF, WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken, Xenopus laevis, Drosophila |
| Conjugate: | Unconjugated |
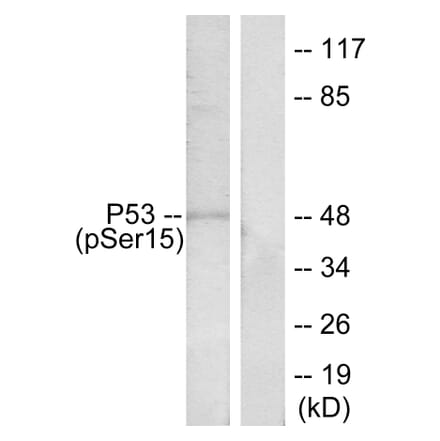
Anti-p53 (phospho Ser15) Antibody (A94113) | |
10µg - 100µg/$190 – $475 | |
| Description: | Rabbit polyclonal antibody to p53 (phospho Ser15). |
| Applications: | WB, IHC, IP, ELISA |
| Reactivity: | Human, Rat |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [rGROEL/780] - BSA and Azide free (A252069) | |
100µg/$545 | |
| Description: | Recombinant mouse monoclonal [rGROEL/780] antibody to HSP60. |
| Applications: | WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken, Xenopus laevis, Drosophila |
| Conjugate: | Unconjugated |
Anti-PHB Antibody (A32316) | |
50µl - 100µl/$455 – $635 | |
| Description: | Rabbit monoclonal antibody to PHB |
| Applications: | WB, ICC, IHC, IP, FC |
| Reactivity: | Human, Mouse, Rat, Zebrafish |
| Conjugate: | Unconjugated |
Anti-HIF-1alpha Antibody (A95079) | |
10µg - 100µg/$190 – $475 | |
| Description: | Rabbit polyclonal antibody to HIF-1alpha. |
| Applications: | WB, IHC, ELISA |
| Reactivity: | Human, Mouse, Rat |
| Conjugate: | Unconjugated |
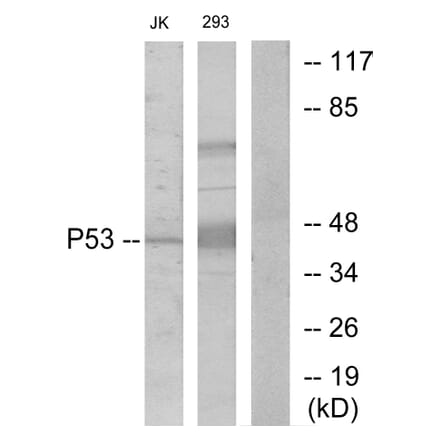
Anti-p53 Antibody (A100048) | |
10µg - 100µg/$190 – $475 | |
| Description: | Rabbit polyclonal antibody to p53. |
| Applications: | WB, IHC, ELISA |
| Reactivity: | Human |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [rGROEL/780] (A248889) | |
100µg/$545 | |
| Description: | Recombinant mouse monoclonal [rGROEL/780] antibody to HSP60. |
| Applications: | WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken, Xenopus laevis, Drosophila |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [HSPD1/2206R] (A248893) | |
100µg/$545 | |
| Description: | Recombinant rabbit monoclonal [HSPD1/2206R] antibody to HSP60. |
| Applications: | WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [HSPD1/2206R] - BSA and Azide free (A252073) | |
100µg/$545 | |
| Description: | Recombinant rabbit monoclonal [HSPD1/2206R] antibody to HSP60. |
| Applications: | WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken |
| Conjugate: | Unconjugated |
Anti-HSP70 Antibody [3A3] (A305077) | |
100µg/$485 | |
| Description: | Mouse monoclonal [3A3] antibody to HSP70. |
| Applications: | WB, IHC, ICC/IF, IP, Antibody Microarray |
| Reactivity: | Human, Mouse, Rat, Amphibians, Chicken, Fish, Yeast, Drosophila, Bacteria, Brine Shrimp |
| Conjugate: | Unconjugated |
Anti-p53 Antibody (A95477) | |
10µg - 100µg/$190 – $475 | |
| Description: | Rabbit polyclonal antibody to p53. |
| Applications: | WB, IHC, IF, ELISA |
| Reactivity: | Human, Mouse, Rat |
| Conjugate: | Unconjugated |
Anti-Tau Antibody [5B10] (A85415) | |
10µl - 100µl/$190 – $545 | |
| Description: | Mouse monoclonal [5B10] antibody to Tau. |
| Applications: | WB, ICC/IF, IHC |
| Reactivity: | Human, Rat, Mouse |
| Conjugate: | Unconjugated |
Anti-Hsp70 Antibody (A32267) | |
50µl - 100µl/$455 – $635 | |
| Description: | Rabbit monoclonal antibody to Hsp70 |
| Applications: | WB, ICC/IF, IHC, FC |
| Reactivity: | Human, Mouse, Rat |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [rHSPD1/6497] (A277894) | |
100µg/$545 | |
| Description: | Recombinant mouse monoclonal [rHSPD1/6497] antibody to HSP60. |
| Applications: | WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [rHSPD1/6497] - BSA and Azide free (A278482) | |
100µg/$545 | |
| Description: | Recombinant mouse monoclonal [rHSPD1/6497] antibody to HSP60. |
| Applications: | WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [HSPD1/6498R] - BSA and Azide free (A278586) | |
100µg/$545 | |
| Description: | Recombinant rabbit monoclonal [HSPD1/6498R] antibody to HSP60. |
| Applications: | WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody (A305092) | |
100µg/$470 | |
| Description: | Rabbit polyclonal antibody to HSP60. |
| Applications: | WB, IHC, IP, ELISA |
| Reactivity: | Human, Mouse, Rat, Bovine, Canine, Chicken, Rabbit, Hamster |
| Conjugate: | Unconjugated |
Anti-Tau (phospho Ser202 + Thr205) Antibody [AH36] (A304919) | |
100µg/$725 | |
| Description: | Rabbit monoclonal [AH36] antibody to Tau (phospho Ser202 + Thr205). |
| Applications: | WB, Dot, ICC/IF, ELISA, IHC |
| Reactivity: | Human, Mouse |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [HSPD1/6498R] (A277998) | |
100µg/$545 | |
| Description: | Recombinant rabbit monoclonal [HSPD1/6498R] antibody to HSP60. |
| Applications: | WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken |
| Conjugate: | Unconjugated |
Anti-HSP70 Antibody [5A5] (A305076) | |
100µg/$485 | |
| Description: | Mouse monoclonal [5A5] antibody to HSP70. |
| Applications: | WB, IHC, ICC/IF, IP, Antibody Microarray |
| Reactivity: | Human, Mouse, Rat, Amphibians, Chicken, Fish, Yeast, Drosophila |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [HSPD1/6496R] (A248892) | |
100µg/$545 | |
| Description: | Recombinant rabbit monoclonal [HSPD1/6496R] antibody to HSP60. |
| Applications: | WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken |
| Conjugate: | Unconjugated |
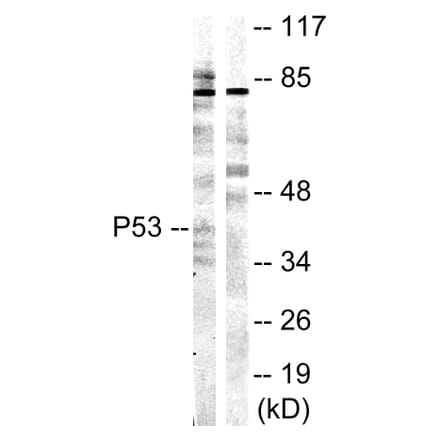
Anti-p53 Antibody [rBP53-12] (A250176) | |
100µg/$5451 Citation | |
| Description: | Recombinant mouse monoclonal [rBP53-12] antibody to p53. |
| Applications: | WB, IHC-P |
| Reactivity: | Human, Monkey, Canine, Hamster, Chicken |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody [HSPD1/6496R] - BSA and Azide free (A252072) | |
100µg/$545 | |
| Description: | Recombinant rabbit monoclonal [HSPD1/6496R] antibody to HSP60. |
| Applications: | WB, IHC-P |
| Reactivity: | Human, Mouse, Rat, Hamster, Sheep, Rabbit, Bovine, Canine, Porcine, Monkey, Chicken |
| Conjugate: | Unconjugated |
Anti-HSP70 Antibody [2A4] (A305075) | |
100µg/$485 | |
| Description: | Mouse monoclonal [2A4] antibody to HSP70. |
| Applications: | WB, IHC, ICC/IF, IP, Antibody Microarray |
| Reactivity: | Human, Mouse, Rat, Amphibians, Chicken, Fish, Yeast, Drosophila |
| Conjugate: | Unconjugated |
Anti-AIF Antibody (A82697) | |
100µg/$495 | |
| Description: | Goat polyclonal antibody to AIF. |
| Applications: | ELISA, WB, IHC, IF |
| Reactivity: | Human, Mouse, Rat, Porcine |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody (A95220) | |
10µg - 100µg/$190 – $475 | |
| Description: | Rabbit polyclonal antibody to HSP60. |
| Applications: | WB, IHC, IF, ELISA |
| Reactivity: | Human, Mouse, Rat |
| Conjugate: | Unconjugated |
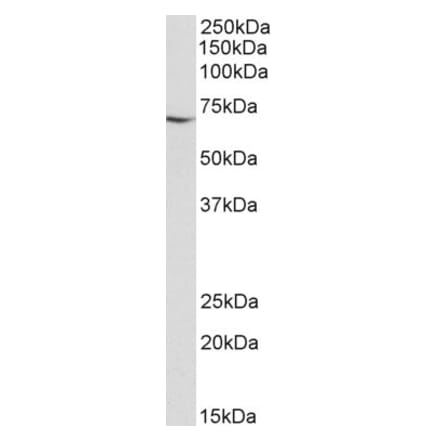
Anti-HSP60 Antibody [HSPD1/875] (A248888) | |
100µg/$545 | |
| Description: | Mouse monoclonal [HSPD1/875] antibody to HSP60. |
| Applications: | IF, WB, IHC-P |
| Reactivity: | Human, Monkey, Mouse, Rat, Hamster, Guinea Pig, Chicken |
| Conjugate: | Unconjugated |
Anti-p53 Antibody (A295374) | |
100µl/$490 | |
| Description: | Rabbit polyclonal antibody to p53. |
| Applications: | WB, IP, IHC, ChIP-Seq, Flow Cytometry |
| Reactivity: | Human |
| Conjugate: | Unconjugated |
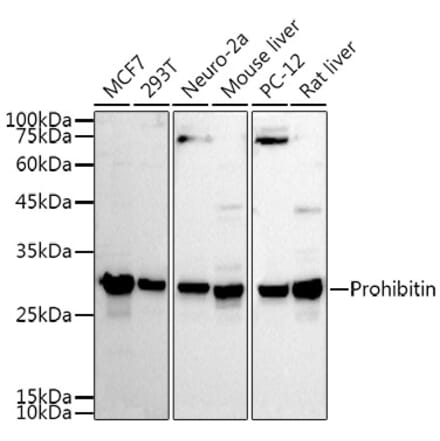
Anti-Prohibitin Antibody (A12508) | |
100µl/$355 | |
| Description: | Rabbit polyclonal antibody to Prohibitin. |
| Applications: | WB, IHC, ICC/IF |
| Reactivity: | Human, Mouse, Rat |
| Conjugate: | Unconjugated |
Anti-HSP60 Antibody (A12728) | |
100µl/$355 | |
| Description: | Rabbit polyclonal antibody to HSP60. |
| Applications: | WB, IHC, ICC/IF, IP |
| Reactivity: | Human, Mouse, Rat |
| Conjugate: | Unconjugated |
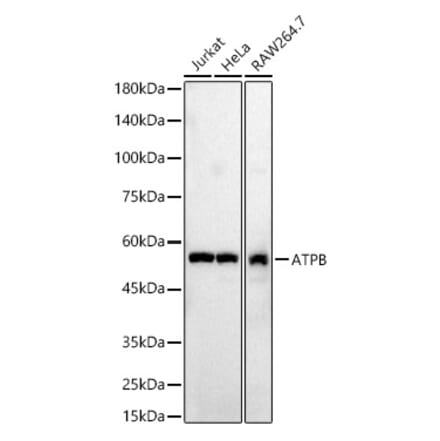
Anti-ATPB Antibody (A14964) | |
100µl/$355 | |
| Description: | Rabbit polyclonal antibody to ATPB. |
| Applications: | WB, IHC, ICC/IF |
| Reactivity: | Human, Mouse, Rat |
| Conjugate: | Unconjugated |
Showing 1-50 of 694 products
![Immunofluorescence - Anti-Tau Antibody [2E9] (A85416) - Antibodies.com](https://cdn.antibodies.com/image/catalog/85/A85416_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP70 Antibody [BB70] (A305113) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305113_1.png?profile=product_search)
![Western Blot - Anti-HSP70 Antibody [N27F3-4] (A305087) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305087_1.png?profile=product_search)
![Immunofluorescence - Anti-HSP60 Antibody [LK2] - BSA and Azide free (A252064) - Antibodies.com](https://cdn.antibodies.com/image/catalog/252/A252065_1.jpg?profile=product_search)
![Immunofluorescence - Anti-HSP60 Antibody [LK2] (A248884) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248885_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [LK2] (A305090) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305090_1.png?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [LK1] - BSA and Azide free (A252063) - Antibodies.com](https://cdn.antibodies.com/image/catalog/252/A252063_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [HSPD1/780] - BSA and Azide free (A252067) - Antibodies.com](https://cdn.antibodies.com/image/catalog/252/A252067_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [LK1] (A248883) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248883_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [HSPD1/780] (A248887) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248887_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [SPM253] - BSA and Azide free (A252064) - Antibodies.com](https://cdn.antibodies.com/image/catalog/252/A252064_1.jpg?profile=product_search)

![Immunohistochemistry - Anti-HSP60 Antibody [SPM253] (A248884) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248884_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [rGROEL/780] - BSA and Azide free (A252069) - Antibodies.com](https://cdn.antibodies.com/image/catalog/252/A252069_1.jpg?profile=product_search)


![Immunohistochemistry - Anti-HSP60 Antibody [rGROEL/780] (A248889) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248889_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [HSPD1/2206R] (A248893) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248893_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [HSPD1/2206R] - BSA and Azide free (A252073) - Antibodies.com](https://cdn.antibodies.com/image/catalog/252/A252073_1.jpg?profile=product_search)
![Western Blot - Anti-HSP70 Antibody [3A3] (A305077) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305077_1.png?profile=product_search)
![Immunofluorescence - Anti-Tau Antibody [5B10] (A85415) - Antibodies.com](https://cdn.antibodies.com/image/catalog/85/A85415_1.jpg?profile=product_search)

![Immunohistochemistry - Anti-HSP60 Antibody [rHSPD1/6497] (A277894) - Antibodies.com](https://cdn.antibodies.com/image/catalog/277/A277894_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [rHSPD1/6497] - BSA and Azide free (A278482) - Antibodies.com](https://cdn.antibodies.com/image/catalog/278/A278482_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [HSPD1/6498R] - BSA and Azide free (A278586) - Antibodies.com](https://cdn.antibodies.com/image/catalog/278/A278586_1.jpg?profile=product_search)

![Immunohistochemistry - Anti-Tau (phospho Ser202 + Thr205) Antibody [AH36] (A304919) - Antibodies.com](https://cdn.antibodies.com/image/catalog/304/A304919_1.png?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [HSPD1/6498R] (A277998) - Antibodies.com](https://cdn.antibodies.com/image/catalog/277/A277998_1.jpg?profile=product_search)
![Western Blot - Anti-HSP70 Antibody [5A5] (A305076) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305076_1.png?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [HSPD1/6496R] (A248891) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248892_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-p53 Antibody [rBP53-12] (A250176) - Antibodies.com](https://cdn.antibodies.com/image/catalog/250/A250176_1.jpg?profile=product_search)
![Immunohistochemistry - Anti-HSP60 Antibody [HSPD1/6496R] - BSA and Azide free (A252071) - Antibodies.com](https://cdn.antibodies.com/image/catalog/252/A252072_1.jpg?profile=product_search)
![Western Blot - Anti-HSP70 Antibody [2A4] (A305075) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305075_1.png?profile=product_search)

![Immunofluorescence - Anti-HSP60 Antibody [HSPD1/875] (A248888) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248888_1.jpg?profile=product_search)