Discover our catalog of 21 highly specific SIRT1 antibodies validated for Western Blot, Western Blotting, Western Blotting (WB), Immunohistochemistry, Immunocytochemistry and more. Our product range includes monoclonal and polyclonal antibodies reactive with samples from human, mouse, rat, monkey and chicken. SIRT1 antibodies are commonly used as nuclear markers. These antibodies are available in carrier free formulations, unconjugated or directly conjugated to Biotin. Some antibodies have also been validated by protein arrays to confirm their specificity. Several antibodies are also available in trial sizes.
Synonyms: hSIR2, hSIRT1, NAD-dependent protein deacetylase sirtuin-1, NAD-dependent protein deacylase sirtuin-1, Regulatory protein SIR2 homolog 1, SIR2-like protein 1 and SIR2L1
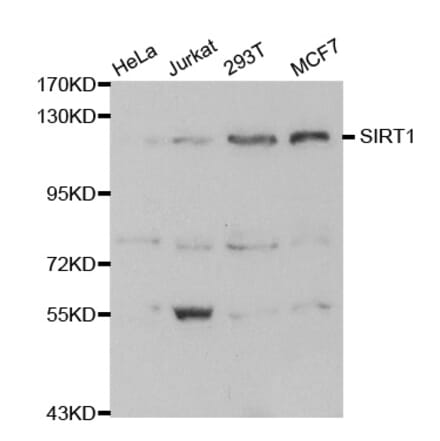
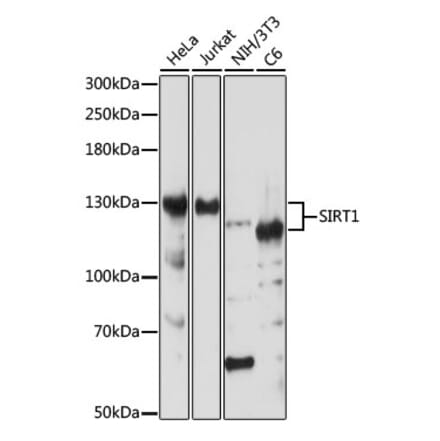
Anti-SIRT1 Antibody (A91756) | |
100µl/$345 | |
| Description: | Rabbit polyclonal antibody to SIRT1. |
| Applications: | WB, ICC/IF, IP, ChIP |
| Reactivity: | Human, Mouse |
| Conjugate: | Unconjugated |
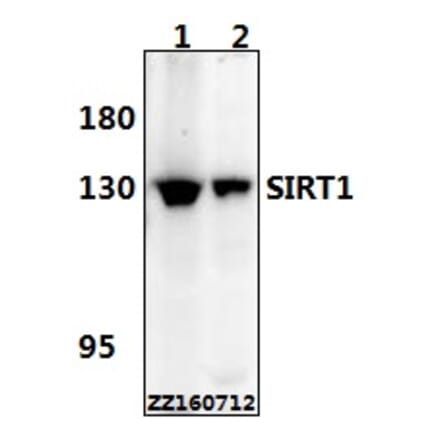
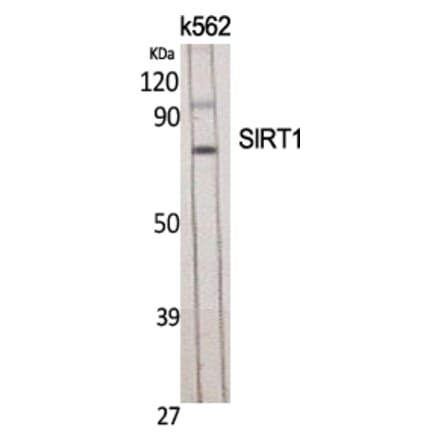
Anti-SirT1 Antibody (A97227) | |
10µg - 100µg/$190 – $475 | |
| Description: | Rabbit polyclonal antibody to SirT1. |
| Applications: | WB, IHC, ELISA |
| Reactivity: | Human, Mouse |
| Conjugate: | Unconjugated |
Anti-SIRT1 Antibody [PCRP-SIRT1-1E11] (A277616) | |
100µg/$545 | |
| Description: | Mouse monoclonal [PCRP-SIRT1-1E11] antibody to SIRT1. |
| Applications: | ELISA, IP, Flow Cytometry, IF, WB |
| Reactivity: | Human |
| Conjugate: | Unconjugated |
![Flow Cytometry - Anti-SIRT1 Antibody [PCRP-SIRT1-1E11] (A277616) - Antibodies.com](https://cdn.antibodies.com/image/catalog/277/A277616_1.jpg?profile=product_search)
Anti-SIRT1 Antibody [PCRP-SIRT1-1E11] - BSA and Azide free (A278204) | |
100µg/$545 | |
| Description: | Mouse monoclonal [PCRP-SIRT1-1E11] antibody to SIRT1. |
| Applications: | ELISA, IP, Flow Cytometry, IF, WB |
| Reactivity: | Human |
| Conjugate: | Unconjugated |
![Flow Cytometry - Anti-SIRT1 Antibody [PCRP-SIRT1-1E11] - BSA and Azide free (A278204) - Antibodies.com](https://cdn.antibodies.com/image/catalog/278/A278204_1.jpg?profile=product_search)
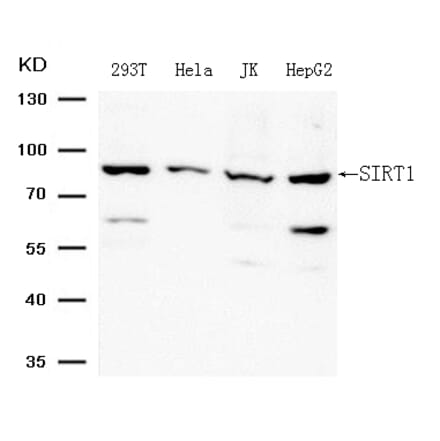
Anti-SIRT1 Antibody (A80509) | |
100µl/$345 | |
| Description: | Rabbit polyclonal antibody to SIRT1. |
| Applications: | WB, IHC |
| Reactivity: | Human, Mouse |
| Conjugate: | Unconjugated |
Anti-SIRT1 Antibody (A82815) | |
100µg/$495 | |
| Description: | Goat polyclonal antibody to SIRT1. |
| Applications: | ELISA, WB, IHC |
| Reactivity: | Human |
| Conjugate: | Unconjugated |
Anti-SirT1 Antibody (A97228) | |
10µg - 100µg/$190 – $475 | |
| Description: | Rabbit polyclonal antibody to SirT1. |
| Applications: | WB, IHC, ELISA |
| Reactivity: | Human, Mouse |
| Conjugate: | Unconjugated |
Anti-SIRT1 Antibody (A12558) | |
100µl/$345 | |
| Description: | Rabbit polyclonal antibody to SIRT1. |
| Applications: | WB, ICC/IF |
| Reactivity: | Human, Mouse, Rat |
| Conjugate: | Unconjugated |
Anti-SIRT1 Antibody (A84243) | |
100µg/$495 | |
| Description: | Goat polyclonal antibody to SIRT1. |
| Applications: | ELISA, WB |
| Reactivity: | Mouse |
| Conjugate: | Unconjugated |
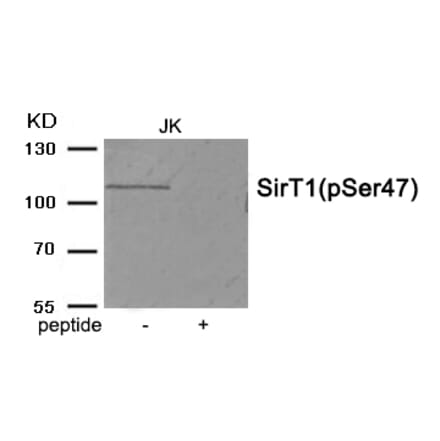
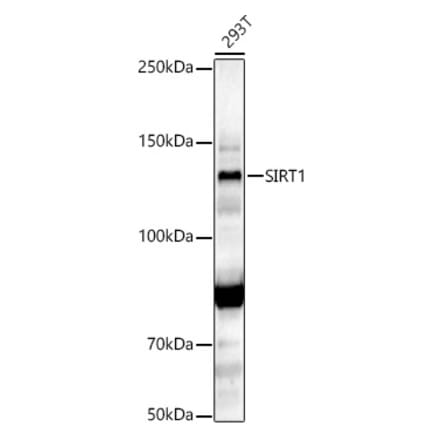
Anti-SIRT1 (phospho Ser47) Antibody [ARC1520] (A305791) | |
100µl/$560 | |
| Description: | Rabbit monoclonal [ARC1520] antibody to SIRT1 (phospho Ser47). |
| Applications: | WB |
| Reactivity: | Human, Mouse |
| Conjugate: | Unconjugated |
Anti-SIRT1 Antibody (A308900) | |
100µl/$345 | |
| Description: | Rabbit polyclonal antibody to SIRT1. |
| Applications: | WB |
| Reactivity: | Human |
| Conjugate: | Unconjugated |
Anti-SIRT1 Antibody (A283427) | |
100µg/$435 | |
| Description: | Rabbit polyclonal antibody to SIRT1. |
| Applications: | WB |
| Reactivity: | Human |
| Conjugate: | Unconjugated |
Anti-SIRT1 Antibody (A54916) | |
100µg/$485 | |
| Description: | Rabbit polyclonal antibody to SIRT1 |
| Applications: | ELISA, IHC, IP, WB |
| Reactivity: | Chicken, Human, Monkey, Mouse, Rat |
| Conjugate: | Unconjugated |
Anti-SIRT1 Antibody (A34201) | |
50µl - 100µl/$390 – $560 | |
| Description: | Rabbit polyclonal antibody to SIRT1 |
| Applications: | WB, IHC, IF, IP, CHIP |
| Reactivity: | Human |
| Conjugate: | Unconjugated |
Anti-SIRT1 Antibody (Biotin) (A53237) | |
100µg/$560 | |
| Description: | Rabbit polyclonal antibody to SIRT1 (Biotin) |
| Applications: | ELISA, IHC, IP, WB |
| Reactivity: | Chicken, Human, Monkey, Mouse, Rat |
| Conjugate: | Biotin |
Anti-SIRT1 Antibody (A29757) | |
50µl - 100µl/$435 – $635 | |
| Description: | Rabbit polyclonal antibody to SIRT1 |
| Applications: | WB, IHC, IF, IP, CHIP |
| Reactivity: | Human, Rat |
| Conjugate: | Unconjugated |
Anti-SIRT1 Antibody (FITC) (A54102) | |
100µg/$560 | |
| Description: | Rabbit polyclonal antibody to SIRT1 (FITC) |
| Applications: | ELISA, IHC, IP, WB |
| Reactivity: | Chicken, Human, Monkey, Mouse, Rat |
| Conjugate: | FITC |
Anti-SIRT1 Antibody (A48660) | |
100µl/$735 | |
| Description: | Chicken polyclonal antibody to SIRT1 |
| Applications: | E, WB, IF |
| Reactivity: | Human, Mouse, Rat |
| Conjugate: | Unconjugated |
Anti-SIRT1 Antibody (A35964) | |
50µl - 100µl/$275 – $405 | |
| Description: | Rabbit polyclonal antibody to SIRT1 |
| Applications: | WB, IHC, ELISA |
| Reactivity: | Human, Mouse |
| Conjugate: | Unconjugated |
Anti-SIRT1 (Ab-344) Antibody (A41321) | |
50µl - 100µl/$275 – $405 | |
| Description: | Rabbit polyclonal antibody to SIRT1 (Ab-344) |
| Applications: | WB |
| Reactivity: | Human, Mouse, Rat |
| Conjugate: | Unconjugated |
Anti-SirT1 (Phospho-Ser47) Antibody (A51159) | |
50µl - 100µl/$275 – $405 | |
| Description: | Rabbit polyclonal antibody to SirT1 (Phospho-Ser47) |
| Applications: | WB |
| Reactivity: | Human |
| Conjugate: | Unconjugated |
Showing 1-21 of 21 products
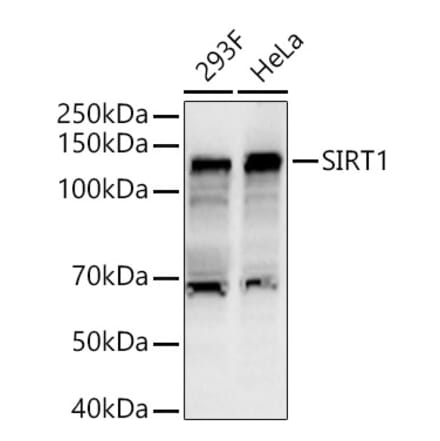

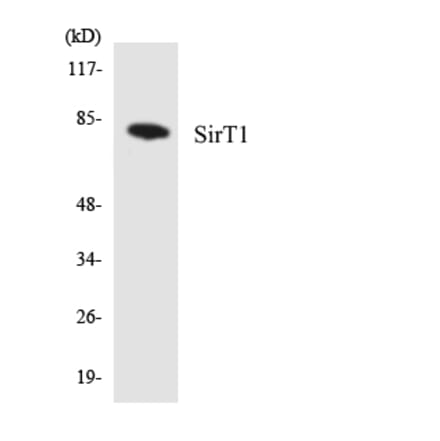

![Western Blot - Anti-SIRT1 (phospho Ser47) Antibody [ARC1520] (A305791) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305791_1.jpg?profile=product_search)