Unconjugated
CD98hc (SLC3A2) constitutively and specifically associates with beta(1) integrins and is highly expressed on the surface of human tumor cells irrespective of the tissue of origin. We have found here that expression of CD98hc promotes both anchorage- and serum-independent growth. This oncogenic activity is dependent on beta(1) integrin-mediated phosphoinositol 3-hydroxykinase stimulation and the level of surface expression of CD98hc. Using chimeras of CD98hc and the type II membrane protein CD69, we show that the transmembrane domain of CD98hc is necessary and sufficient for integrin association in cells. Furthermore, CD98hc/beta(1) integrin association is required for focal adhesion kinase-dependent phosphoinositol 3-hydroxykinase activation and cellular transformation. Amino acids 82-87 in the putative cytoplasmic/transmembrane region appear to be critical for the oncogenic potential of CD98hc and provide a novel mechanism for tumor promotion by integrins. These results explain how high expression of CD98hc in human cancers contributes to transformation; furthermore, the transmembrane association of CD98hc and beta(1) integrins may provide a new target for cancer therapy.
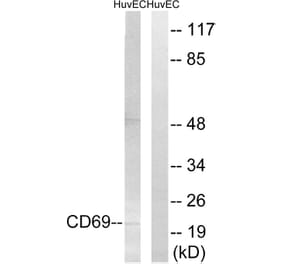
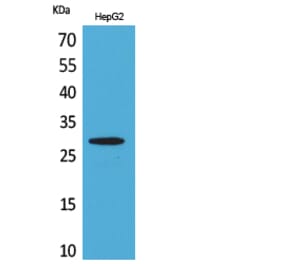
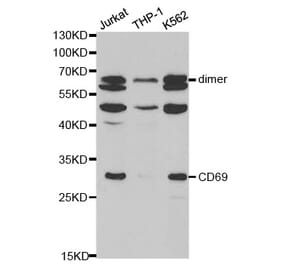
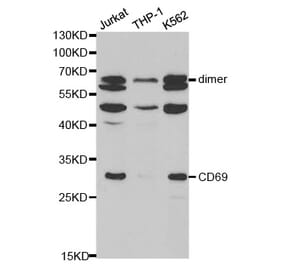
CD69 is expressed on the surface of all hematopoietically derived leukocytes and is suggested to function as a multipurpose cell-surface trigger molecule important in the development and activation of many different cell types. Human CD69 contains only a single consensus sequence for N-linked oligosaccharide addition within its extracellular domain (Asn-Val-Thr), yet exists as two distinct glycoforms that are assembled together into disulfide-linked homodimers and heterodimers. The molecular basis for human CD69 heterogeneity has remained elusive. In the current report we show that human CD69 glycoforms are generated before the egress of CD69 proteins from the endoplasmic reticulum to the Golgi and are synthesized under conditions where Golgi processing is inhibited, effectively ruling out the possibility that CD69 heterogeneity results from the differential processing of a single glycosylation site in the Golgi complex. Importantly, these data demonstrate that contrary to current belief, not one but two sites for N-glycan addition exist within the human CD69 extracellular domain and identify the second, "cryptic" CD69 N-glycan attachment site as the atypical Cys-containing glycosylation motif, Asn-Ala-Cys. The results in this study provide a molecular basis for human CD69 heterogeneity and show that multiple dimeric forms of human CD69 result from the variable addition of N-glycans to atypical and typical glycosylation motifs within the CD69 extracellular domain.




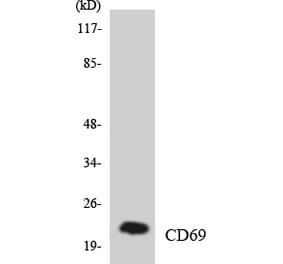
![Western Blot - Anti-CD69 Antibody [ARC56158] (A308834) - Antibodies.com](https://cdn.antibodies.com/image/catalog/308/A308834_1.jpg?profile=product_alternative)
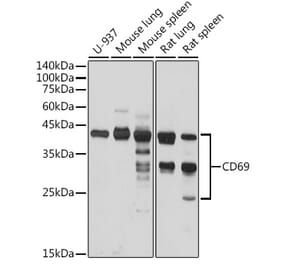
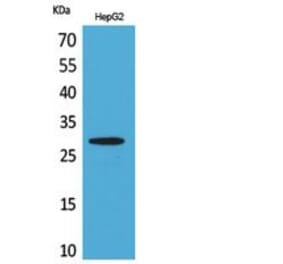
![SDS-PAGE - Anti-CD69 Antibody [Research Grade Biosimilar] - Low endotoxin, Azide free (A323953) - Antibodies.com](https://cdn.antibodies.com/image/catalog/323/A323953_1.jpg?profile=product_alternative)