This antibody MEL-14 reacts with mouse CD62L (L-selectin), a 75 kDa single chain type I glycoprotein expressed on most peripheral blood B lymphocytes, T lymphocytes, monocytes and granulocytes; it is also present on a subset of NK cells and certain hematopoietic malignant cells.
Applications
Flow Cytometry, IP, IHC-P, IHC-Fr, ICC
Dilutions
Flow Cytometry: 1-4 µg/ml.
Reactivity
Mouse
Immunogen
C3H/eb mouse B cell lymphoma 38C-13.
Host
Rat
Clonality
Monoclonal
Clone ID
Mel-14
Isotype
IgG2a
Conjugate
Unconjugated
Purification
Protein G chromatography.
Concentration
1 mg/ml
Predicted MW
75 kDa
Product Form
Liquid
Formulation
Supplied in Phosphate Buffered Saline, pH 7.4, with 15 mM Sodium Azide.
Storage
Shipped at 4°C. Upon delivery aliquot and store at -20°C. Avoid freeze/thaw cycles.




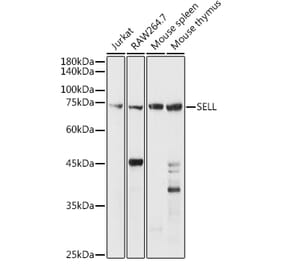

![SDS-PAGE - Anti-CD62L Antibody [LAM1-116] (A249950) - Antibodies.com](https://cdn.antibodies.com/image/catalog/249/A249951_1.jpg?profile=product_alternative)
![SDS-PAGE - Anti-CD62L Antibody [LAM1-116] - BSA and Azide free (A253130) - Antibodies.com](https://cdn.antibodies.com/image/catalog/253/A253131_1.jpg?profile=product_alternative)
![Flow Cytometry - Anti-CD62L Chimeric Antibody [DMC439] - Azide free (A318739) - Antibodies.com](https://cdn.antibodies.com/image/catalog/318/A318739_1.jpg?profile=product_alternative)