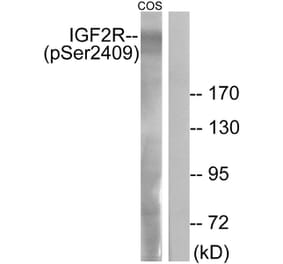
Synonyms
300 kDa mannose 6-phosphate receptor, Cation-independent mannose-6-phosphate receptor, CI Man-6-P receptor, CI-MPR, IGF-II receptor, Insulin-like growth factor 2 receptor, Insulin-like growth factor II receptor, M6P/IGF2 receptor, M6P/IGF2R, M6PR, MPR 300, MPRI


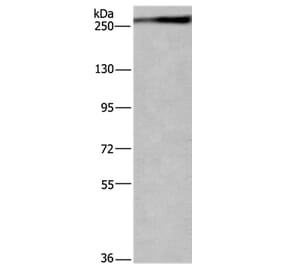
![Western Blot - Anti-CD222 Antibody [MEM-238] (A85552) - Antibodies.com](https://cdn.antibodies.com/image/catalog/85/A85553_59.jpg?profile=product_top)
![Western Blot - Anti-CD222 Antibody [MEM-238] (A85552) - Antibodies.com](https://cdn.antibodies.com/image/catalog/85/A85553_59.jpg?profile=product_top_thumb)
![Western Blot - Anti-CD222 Antibody [MEM-238] (A85552) - Antibodies.com](https://cdn.antibodies.com/image/catalog/85/A85553_59.jpg?profile=product_image)

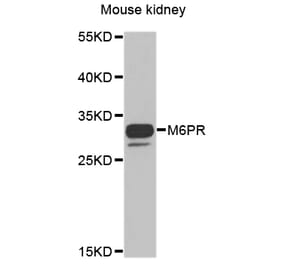
![Western Blot - Anti-M6PR (cation independent) Antibody [ARC0263] (A308985) - Antibodies.com](https://cdn.antibodies.com/image/catalog/308/A308985_1.jpg?profile=product_alternative)
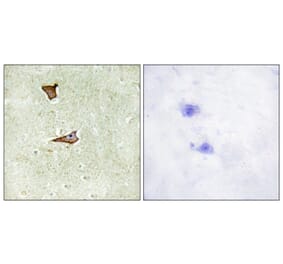
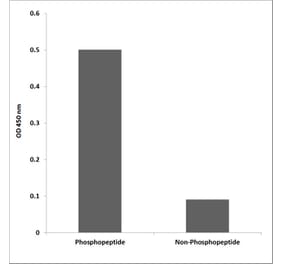
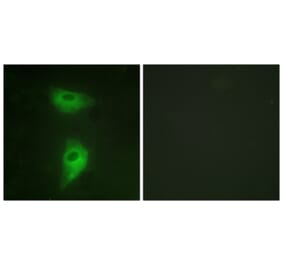
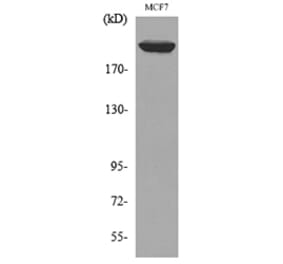
![Flow Cytometry - Anti-CD222 Antibody [MEM-240] (A86835) - Antibodies.com](https://cdn.antibodies.com/image/catalog/86/A86836_921.jpg?profile=product_alternative)