

Unconjugated
Memory formation in the brain is thought to rely on the remodeling of synaptic connections which eventually results in neural network rewiring. This remodeling is likely to involve ultrathin astroglial protrusions which often occur in the immediate vicinity of excitatory synapses. The phenomenology, cellular mechanisms, and causal relationships of such astroglial restructuring remain, however, poorly understood. This is in large part because monitoring and probing of the underpinning molecular machinery on the scale of nanoscopic astroglial compartments remains a challenge. Here we briefly summarize the current knowledge regarding the cellular organisation of astroglia in the synaptic microenvironment and discuss molecular mechanisms potentially involved in use-dependent astroglial morphogenesis. We also discuss recent observations concerning morphological astroglial plasticity, the respective monitoring methods, and some of the newly emerging techniques that might help with conceptual advances in the area.
© 2015 The Authors. Glia Published by Wiley Periodicals, Inc.
Apelin, a novel peptide originally isolated from bovine stomach tissue extracts, is widely but selectively distributed throughout the nervous system. Vasopressin and oxytocin are synthesized in the magnocellular neurons of the hypothalamic supraoptic nucleus (SON) and paraventricular nucleus, which are apelin-rich regions in the central nervous system. We made extracellular electrophysiological recordings from the transpharyngeally exposed SON of urethane-anaesthetized rats to assess the role of apelin in the control of the firing activity of identified magnocellular vasopressin and oxytocin neurons in vivo. Apelin-13 administration onto SON neurons via microdialysis revealed cell-specific responses; apelin-13 increased the firing rates of vasopressin cells but had no effect on the firing rate of oxytocin neurons. A direct excitatory effect of apelin-13 on vasopressin cell activity is also supported by our in vitro studies showing depolarization of membrane potential and increase in action potential firing. To assess the effects of apelin-13 on somatodendritic peptide release, we used in vitro release studies from SON explants in combination with highly sensitive and specific RIA. Apelin-13 decreases basal (by 78%; P < 0.05; n = 6) and potassium-stimulated (by 57%; P < 0.05; n = 6) vasopressin release but had no effect on somatodendritic oxytocin release. Taken together, our data suggest a local autocrine feedback action of apelin on magnocellular vasopressin neurons. Furthermore, these data show a marked dissociation between axonal and dendritic vasopressin release with a decrease in somatodendritic release but an increase in electrical activity at the cell bodies, indicating that release from these two compartments can be regulated wholly independently.


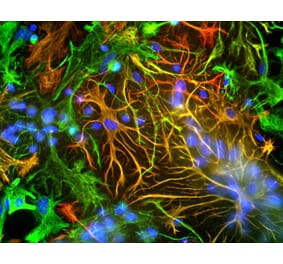
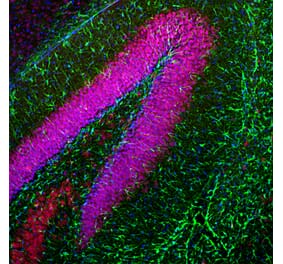
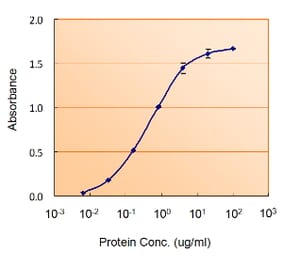
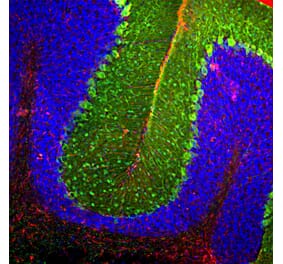




![Immunofluorescence - Anti-GFAP Antibody [5C10] (A85422) - Antibodies.com](https://cdn.antibodies.com/image/catalog/85/A85422_1.jpg?profile=product_alternative)
![Western Blot - Anti-GFAP Antibody [GA-5] - BSA and Azide free (A251887) - Antibodies.com](https://cdn.antibodies.com/image/catalog/251/A251887_1.jpg?profile=product_alternative)
![Western Blot - Anti-GFAP Antibody [GA-5] (A248705) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248705_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-GFAP Antibody [SPM248] - BSA and Azide free (A251888) - Antibodies.com](https://cdn.antibodies.com/image/catalog/251/A251888_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-GFAP Antibody [GA-5 + ASTRO/789] (A248708) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248709_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-GFAP Antibody [GA-5 + ASTRO/789] - BSA and Azide free (A251890) - Antibodies.com](https://cdn.antibodies.com/image/catalog/251/A251891_1.jpg?profile=product_alternative)
![Western Blot - Anti-GFAP Antibody [ARC0206] (A307282) - Antibodies.com](https://cdn.antibodies.com/image/catalog/307/A307282_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-GFAP Antibody [SPM248] (A248706) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248706_1.jpg?profile=product_alternative)

![Immunohistochemistry - Anti-GFAP Antibody [rASTRO/789] (A248711) - Antibodies.com](https://cdn.antibodies.com/image/catalog/248/A248711_1.jpg?profile=product_alternative)