

Unconjugated
The transmembrane chemokines CX3CL1/fractalkine and CXCL16 are widely expressed in different types of tumors, often without an appropriate expression of their classical receptors. We observed that receptor-negative cancer cells could be stimulated by the soluble chemokines. Searching for alternative receptors we detected that all cells expressing or transfected with transmembrane chemokine ligands bound the soluble chemokines with high affinity and responded by phosphorylation of intracellular kinases, enhanced proliferation and anti-apoptosis. This activity requires the intracellular domain and apparently the dimerization of the transmembrane chemokine ligand. Thus, shed soluble chemokines can generate auto- or paracrine signals by binding and activating their transmembrane forms. We term this novel mechanism "inverse signaling". We suppose that inverse signaling is an autocrine feedback and fine-tuning system in the communication between cells that in tumors supports stabilization and proliferation.
BACKGROUND:
In HIV-infected macrophages, newly formed progeny virus particles accumulate in intracellular plasma membrane-connected compartments (IPMCs). Although the virus is usually seen in these compartments, it is unclear whether HIV assembly is specifically targeted to IPMCs or whether some viruses may also form at the cell surface but are not detected, as particles budding from the latter site will be released into the medium.
RESULTS:
To investigate the fidelity of HIV-1 targeting to IPMCs compared to the cell surface directly, we generated mutants defective in recruitment of the Endosomal Sorting Complexes Required for Transport (ESCRT) proteins required for virus scission. For mutants unable to bind the ESCRT-I component Tsg101, HIV release was inhibited and light and electron microscopy revealed that budding was arrested. When expressed in human monocyte-derived macrophages (MDM), these mutants formed budding-arrested, immature particles at their assembly sites, allowing us to capture virtually all of the virus budding events. A detailed morphological analysis of the distribution of the arrested viruses by immunofluorescence staining and confocal microscopy, and by electron microscopy, demonstrated that HIV assembly in MDMs is targeted primarily to IPMCs, with fewer than 5 % of budding events seen at the cell surface. Morphometric analysis of the relative membrane areas at the cell surface and IPMCs confirmed a large enrichment of virus assembly events in IPMCs. Serial block-face scanning electron microscopy of macrophages infected with a budding-defective HIV mutant revealed high-resolution 3D views of the complex organisation of IPMCs, with in excess of 15,000 associated HIV budding sites, and multiple connections between IPMCs and the cell surface.
CONCLUSIONS:
Using detailed quantitative analysis, we demonstrate that HIV assembly in MDMs is specifically targeted to IPMCs. Furthermore, 3D analysis shows, for the first time, the detailed ultrastructure of an IPMC within a large cell volume, at a resolution that allowed identification of individual virus assembly events, and potential portals through which virus may be released during cell-cell transfer. These studies provide new insights to the organisation of the HIV assembly compartments in macrophages, and show how HIV particles accumulating in these protected sites may function as a virus reservoir.
![Flow Cytometry - Anti-CD44 Antibody [MEM-85] (A85615) - Antibodies.com](https://cdn.antibodies.com/image/catalog/85/A85618_109.jpg?profile=product_top)
![Flow Cytometry - Anti-CD44 Antibody [MEM-85] (A85615) - Antibodies.com](https://cdn.antibodies.com/image/catalog/85/A85618_109.jpg?profile=product_top_thumb)
![Flow Cytometry - Anti-CD44 Antibody [MEM-85] (A85615) - Antibodies.com](https://cdn.antibodies.com/image/catalog/85/A85618_109.jpg?profile=product_image)























![Western Blot - Anti-CD44 Antibody [MEM-263] (A85673) - Antibodies.com](https://cdn.antibodies.com/image/catalog/85/A85675_150.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-CD44 Antibody [156-3C11] (A250711) - Antibodies.com](https://cdn.antibodies.com/image/catalog/250/A250711_1.jpg?profile=product_alternative)
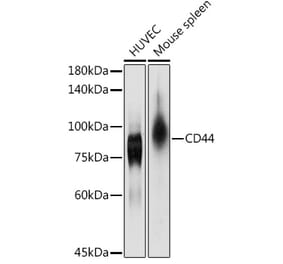
![Immunohistochemistry - Anti-CD44 Antibody [156-3C11] - BSA and Azide free (A253891) - Antibodies.com](https://cdn.antibodies.com/image/catalog/253/A253891_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-CD44 Antibody [HCAM/918] - BSA and Azide free (A253896) - Antibodies.com](https://cdn.antibodies.com/image/catalog/253/A253896_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-CD44 Antibody [rHCAM/918] (A250713) - Antibodies.com](https://cdn.antibodies.com/image/catalog/250/A250714_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-CD44 Antibody [rHCAM/918] - BSA and Azide free (A253893) - Antibodies.com](https://cdn.antibodies.com/image/catalog/253/A253894_1.jpg?profile=product_alternative)
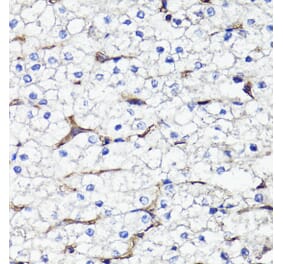
![Immunohistochemistry - Anti-CD44 Antibody [HCAM/918] (A250716) - Antibodies.com](https://cdn.antibodies.com/image/catalog/250/A250716_1.jpg?profile=product_alternative)
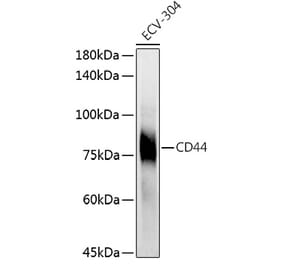

![Immunohistochemistry - Anti-CD44 Antibody [BU75] (A250708) - Antibodies.com](https://cdn.antibodies.com/image/catalog/250/A250709_1.jpg?profile=product_alternative)