Synonyme
DAP1, GAP modifying protein 1, GAP-modifying protein 1, GMP 1, GMP1, OFC10, PIC 1, PIC1, SENP2, Sentrin, Sentrin 1, Small ubiquitin related modifier 1, Small ubiquitin-like modifier 1, Small ubiquitin-related modifier 1, SMT3, SMT3 homolog 3, SMT3 suppressor of mif two 3 homolog 1, SMT3, yeast, homolog 3, Smt3C, SMT3H3, Sumo 1, SUMO-1, SUMO1_HUMAN, Ubiquitin homology domain protein PIC1, Ubiquitin Like 1, Ubiquitin like protein SMT3C, Ubiquitin like protein UBL1, Ubiquitin-homology domain protein PIC1, Ubiquitin-like protein SMT3C, Ubiquitin-like protein UBL1, UBL 1, UBL1


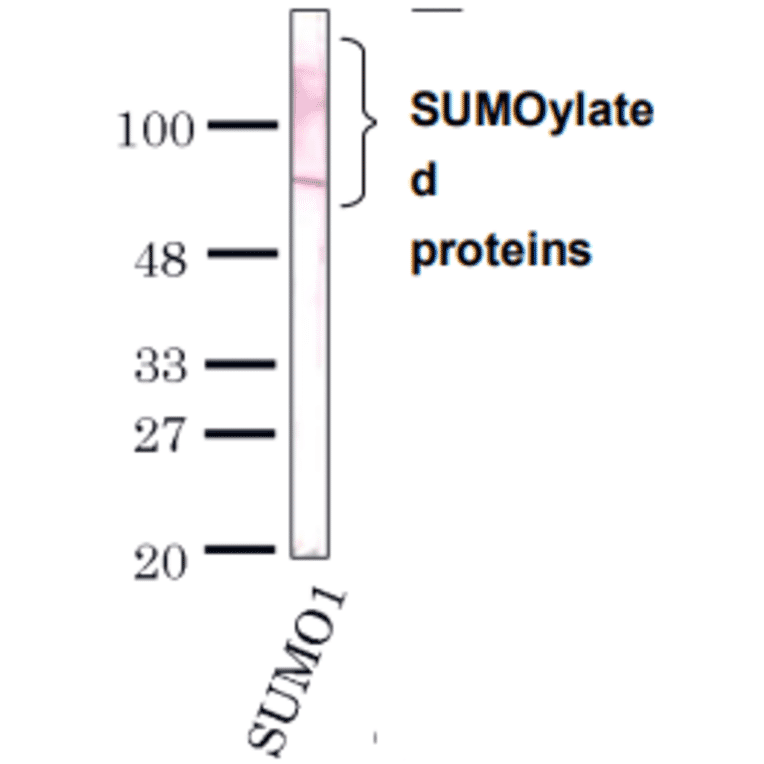
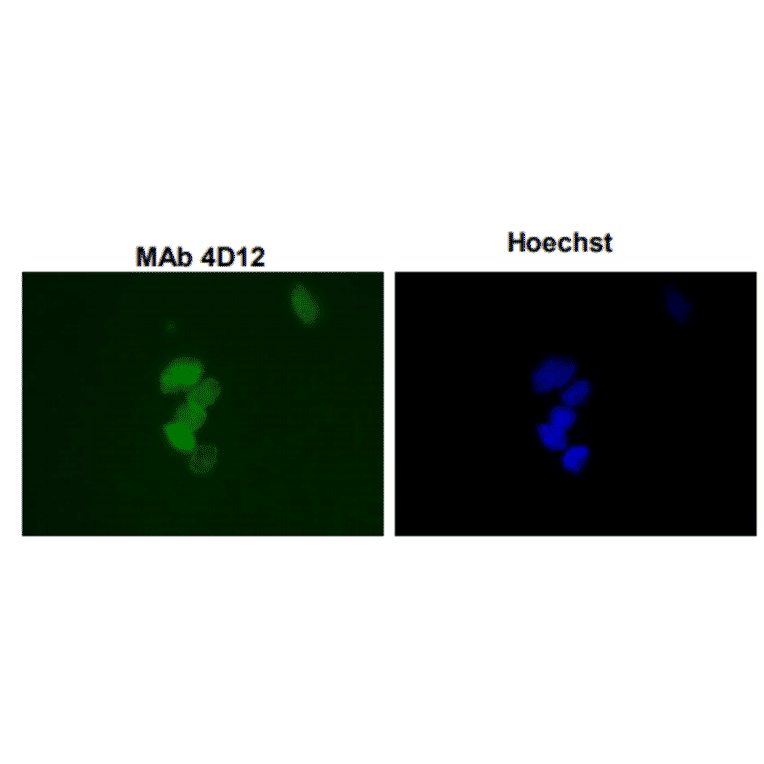
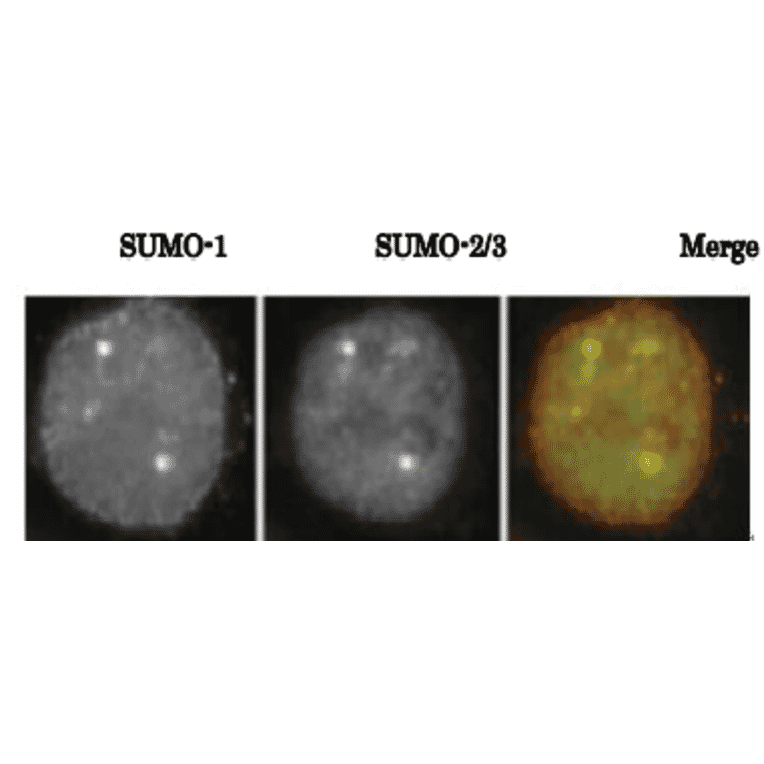
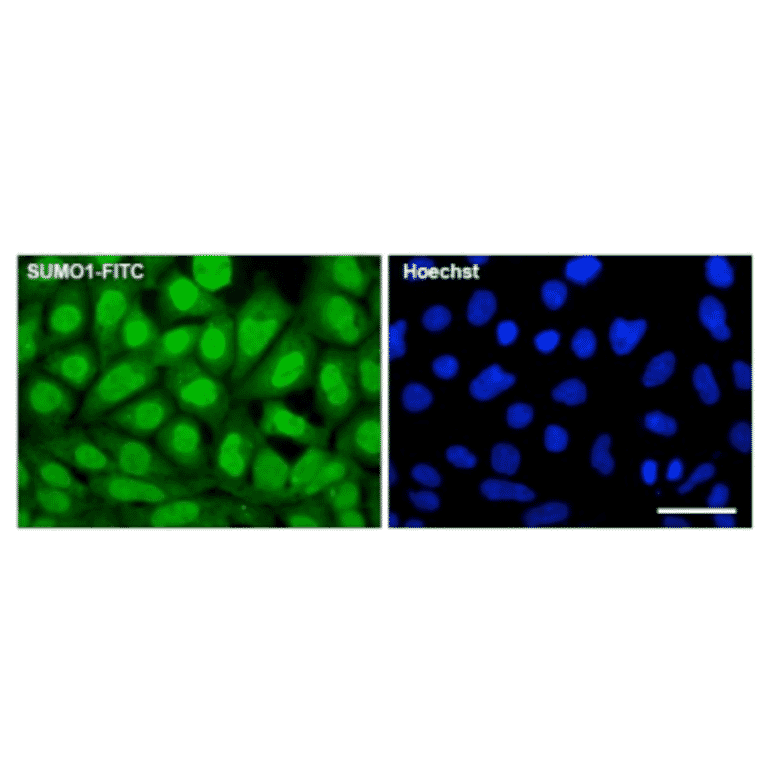
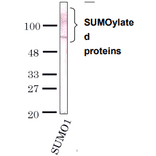
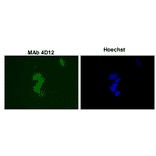
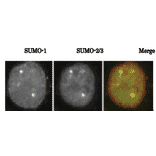
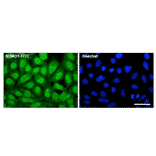

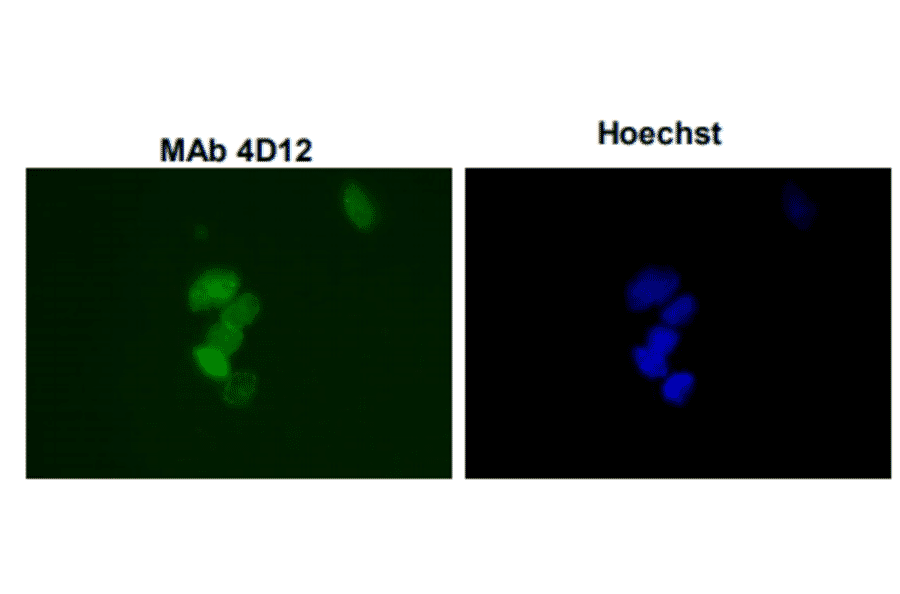
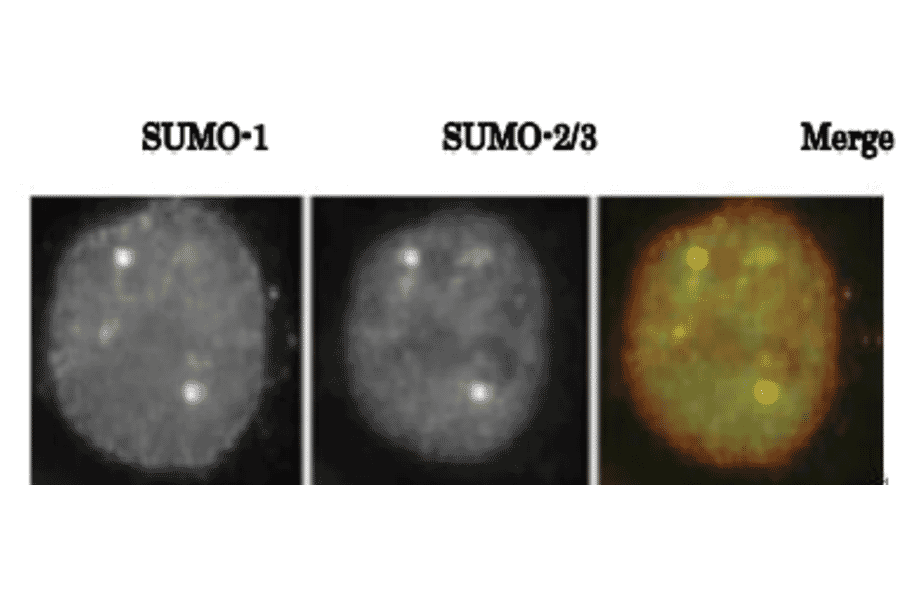
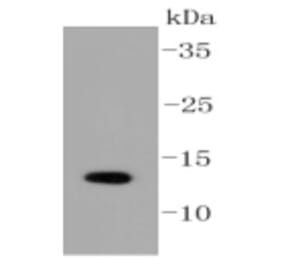
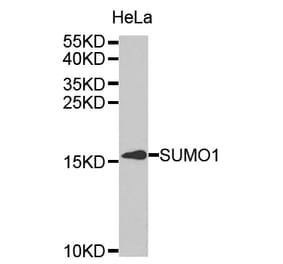
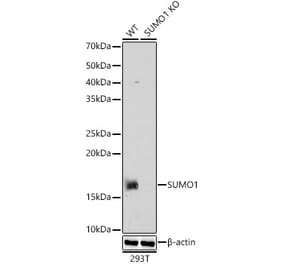
![Western Blot - Anti-Sumo 1 Antibody [ARC0215] (A307733) - Antibodies.com](https://cdn.antibodies.com/image/catalog/307/A307733_1.jpg?profile=product_alternative)
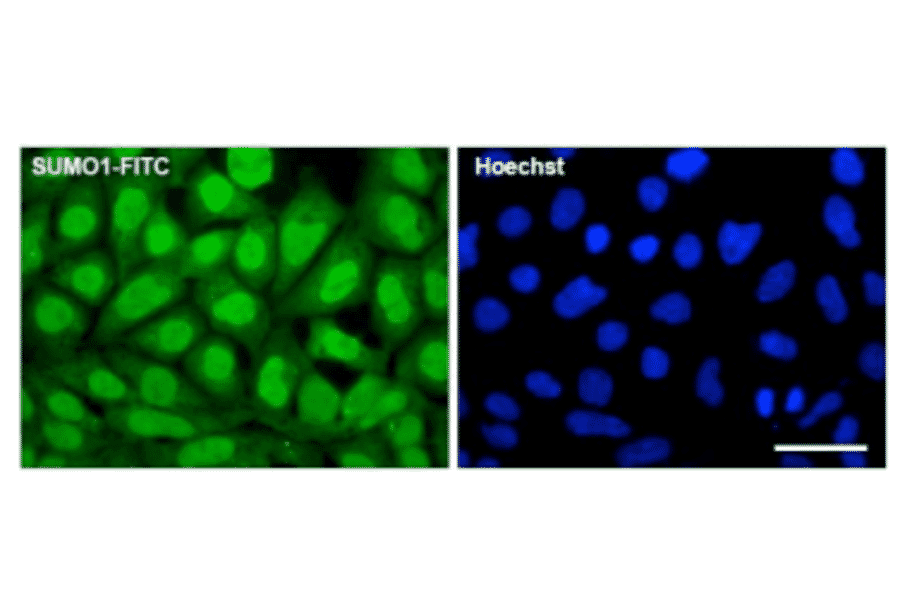
![Immunohistochemistry - Anti-SUMO-1 Antibody [SUMO1/1188] - BSA and Azide free (A253447) - Antibodies.com](https://cdn.antibodies.com/image/catalog/253/A253448_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-SUMO-1 Antibody [SUMO1/1188] (A250267) - Antibodies.com](https://cdn.antibodies.com/image/catalog/250/A250268_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-SUMO-1 Antibody [SM1/495] - BSA and Azide free (A253446) - Antibodies.com](https://cdn.antibodies.com/image/catalog/253/A253446_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-SUMO-1 Antibody [SM1/495] (A250266) - Antibodies.com](https://cdn.antibodies.com/image/catalog/250/A250266_1.jpg?profile=product_alternative)

![Immunohistochemistry - Anti-SUMO-1 Antibody [SPM571] (A250266) - Antibodies.com](https://cdn.antibodies.com/image/catalog/250/A250267_1.jpg?profile=product_alternative)
![Immunohistochemistry - Anti-SUMO-1 Antibody [SPM571] - BSA and Azide free (A253446) - Antibodies.com](https://cdn.antibodies.com/image/catalog/253/A253447_1.jpg?profile=product_alternative)