

Unconjugated
Heat shock factor 1 (HSF1) is a heat shock transcription factor that rapidly induces heat shock gene transcription following thermal stress. In this study, we subjected primary neonatal rat myocardial cells to heat stress in vitro to create a model system for investigating the trends in expression and association between various heat shock proteins (HSPs) and HSF1 under adverse environmental conditions. After the cells were subjected to heat stress at 42°C for different periods of time, HSP and HSF1 mRNA and protein levels were detected by qPCR and western blot analysis in the heat-stressed cells. The HSF1 expression levels significantly increased in the cells following 120 min of exposure to heat stess compared to the levels observed at the beginning of heat stress exposure. HSP90 followed a similar trend in expression to HSF1, whereas HSP70 followed an opposite trend. However, no significant changes were observed in the crystallin, alpha B (CRYAB, also known as HSP beta-5) expression levels during the 480-min period of exposure to heat stress. The interaction between the HSPs and HSF1 was analyzed by STRING 9.1, and it was found that HSF1 interacted with HSP90 and HSP70, and that it did not play a role in regulating CRYAB expression. Based on our findings, HSP70 may suppress HSF1 in rat myocardial cells under conditions of heat stress. Furthermore, our data demonstrate that HSF1 is not the key factor for all HSPs, and this was particularly the case for CRYAB.
Mutations in the C terminus of the serotonin transporter (SERT) disrupt folding and export from the endoplasmic reticulum. Here we examined the hypothesis that a cytosolic heat shock protein relay was recruited to the C terminus to assist folding of SERT. This conjecture was verified by the following observations. (i) The proximal portion of the SERT C terminus conforms to a canonical binding site for DnaK/heat shock protein of 70 kDa (HSP70). A peptide covering this segment stimulated ATPase activity of purified HSP70-1A. (ii) A GST fusion protein comprising the C terminus of SERT pulled down HSP70-1A. The interaction between HSP70-1A and SERT was visualized in live cells by Förster resonance energy transfer: it was restricted to endoplasmic reticulum-resident transporters and enhanced by an inhibitor that traps HSP70-1A in its closed state. (iv) Co-immunoprecipitation confirmed complex formation of SERT with HSP70-1A and HSP90ß. Consistent with an HSP relay, co-chaperones (e.g. HSC70-HSP90-organizing protein) were co-immunoprecipitated with the stalled mutants SERT-R607A/I608A and SERT-P601A/G602A. (v) Depletion of HSP90ß by siRNA or its inhibition increased the cell surface expression of wild type SERT and SERT-F604Q. In contrast, SERT-R607A/I608A and SERT-P601A/G602A were only rendered susceptible to inhibition of HSP70 and HSP90 by concomitant pharmacochaperoning with noribogaine. (vi) In JAR cells, inhibition of HSP90 also increased the levels of SERT, indicating that endogenously expressed transporter was also susceptible to control by HSP90ß. These findings support the concept that the folding trajectory of SERT is sampled by a cytoplasmic chaperone relay.
![Western Blot - Anti-HSP90 alpha Antibody [2G5.G3] (A305031) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305031_1.png?profile=product_top)
![Immunocytochemistry/Immunofluorescence - Anti-HSP90 alpha Antibody [2G5.G3] (A305031) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305031_2.png?profile=product_top)
![Western Blot - Anti-HSP90 alpha Antibody [2G5.G3] (A305031) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305031_1.png?profile=product_top_thumb)
![Immunocytochemistry/Immunofluorescence - Anti-HSP90 alpha Antibody [2G5.G3] (A305031) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305031_2.png?profile=product_top_thumb)
![Western Blot - Anti-HSP90 alpha Antibody [2G5.G3] (A305031) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305031_1.png?profile=product_image)
![Immunocytochemistry/Immunofluorescence - Anti-HSP90 alpha Antibody [2G5.G3] (A305031) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305031_2.png?profile=product_image)









![Immunohistochemistry - Anti-HSP90 alpha Antibody [Hyb-K41009] (A305061) - Antibodies.com](https://cdn.antibodies.com/image/catalog/305/A305061_1.png?profile=product_alternative)

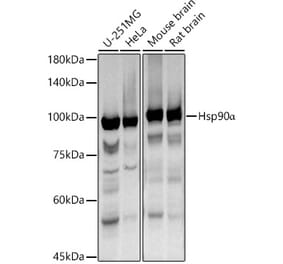
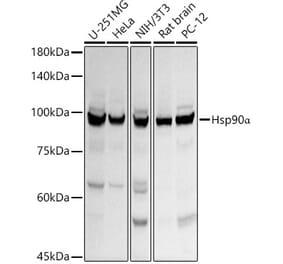
![Western Blot - Anti-Hsp90 alpha Antibody [ARC1167] (A307595) - Antibodies.com](https://cdn.antibodies.com/image/catalog/307/A307595_1.jpg?profile=product_alternative)
![Western Blot - Anti-HSP90 alpha Antibody [M10E3R] (A304896) - Antibodies.com](https://cdn.antibodies.com/image/catalog/304/A304896_1.png?profile=product_alternative)