Unconjugated
Endothelial adhesion is necessary for the hematogenous dissemination of tumor cells. However, the metastatic breast tumor cell MDA-MB-231 does not bind to the endothelium under physiological flow conditions, suggesting alternate mechanisms of adhesion. Since monocytes are highly represented in the tumor microenvironment, and also bind to endothelium during inflammation, we hypothesized that the monocytes assist in the arrest of MDA-MB-231 on the endothelium. Using in vitro models of the dynamic shear environment of the vasculature, we show that TNF-α-activated THP1/primary human monocytes and MDA-MB-231 cells form stable aggregates, and that the monocytes in these aggregates mediate the adhesion of otherwise nonadherent MDA-MB-231 cells to inflamed endothelium under flow (55±2.4 vs. 1.7±0.82 at a shear stress of 0.5 dyn/cm(2), P<0.01). We also show that the hydrodynamic forces determine the size and orientation of aggregates adhered to the endothelium, and strongly favor the attachment of small aggregates with tumor cells downstream of flow (74-86% doublets at 0.5-2 dyn/cm(2), P<0.01). The 5-fold up-regulation of ICAM-1 on TNF-α-activated MDA-MB-231 cells through the Nf-κB pathway was found to be critical in MDA-MB-231-monocyte aggregation and endothelial adhesion. Our results demonstrate that, under inflammatory conditions, monocytes may serve to disseminate tumor cells through circulation, and the tumor-monocyte-endothelial axis may represent a new therapeutic target to reduce cancer metastasis.
The efficient processing of proinsulin into mature insulin and C-peptide is often compromised under conditions of beta cell stress, including diabetes. Impaired proinsulin processing has been challenging to examine by immunofluorescence staining in pancreas tissue because the characterization of antibodies specific for proinsulin, proinsulin intermediates, processed insulin and C-peptide has been limited. This study aimed to identify and characterize antibodies that can be used to detect products of proinsulin processing by immunofluorescence staining in pancreata from different species (mice, rats, dog, pig and human). We took advantage of several knockout mouse lines that lack either an enzyme involved in proinsulin processing or an insulin gene. Briefly, we report antibodies that are specific for several proinsulin processing products, including: a) insulin or proinsulin that has been appropriately processed at the B-C junction; b) proinsulin with a non-processed B-C junction; c) proinsulin with a non-processed A-C junction; d) rodent-specific C-peptide 1; e) rodent-specific C-peptide 2; and f) human-specific C-peptide or proinsulin. In addition, we also describe two 'pan-insulin' antibodies that react with all forms of insulin and proinsulin intermediates, regardless of the species. These antibodies are valuable tools for studying proinsulin processing by immunofluorescence staining and distinguishing between proinsulin products in different species.
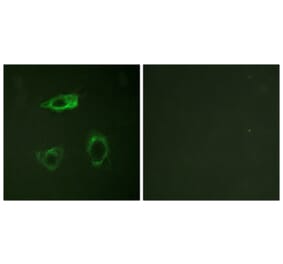
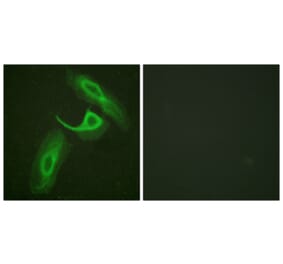
![Immunohistochemistry - Anti-CD18 Antibody [MEM-48] (A86013) - Antibodies.com](https://cdn.antibodies.com/image/catalog/86/A86019_378.jpg?profile=product_top)
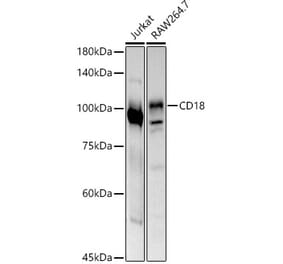
![Western Blot - Anti-CD18 Antibody [MEM-48] (A86019) - Antibodies.com](https://cdn.antibodies.com/image/catalog/86/A86019_379.jpg?profile=product_top)
![Immunohistochemistry - Anti-CD18 Antibody [MEM-48] (A86013) - Antibodies.com](https://cdn.antibodies.com/image/catalog/86/A86019_378.jpg?profile=product_top_thumb)
![Western Blot - Anti-CD18 Antibody [MEM-48] (A86019) - Antibodies.com](https://cdn.antibodies.com/image/catalog/86/A86019_379.jpg?profile=product_top_thumb)
![Immunohistochemistry - Anti-CD18 Antibody [MEM-48] (A86013) - Antibodies.com](https://cdn.antibodies.com/image/catalog/86/A86019_378.jpg?profile=product_image)
![Western Blot - Anti-CD18 Antibody [MEM-48] (A86019) - Antibodies.com](https://cdn.antibodies.com/image/catalog/86/A86019_379.jpg?profile=product_image)



![Western Blot - Anti-CD18 Antibody [MEM-148] (A85610) - Antibodies.com](https://cdn.antibodies.com/image/catalog/85/A85610_101.jpg?profile=product_alternative)